
If enhanced athletic performance and better body composition are important to you I urge you to take notice of the latest in research proven sports nutrition!
To bring some of you up to date, the past year saw the introduction of several research proven legitimate sports nutrition performance enhancing compounds. Namely Citrulline Malate and Beta Alanine paved the way as two of the biggest breakthroughs for 2004. If you are not familiar with these compounds, please read the articles listed here:
- Citrulline Malate: The Next Big Supplement!
- More On Citrulline Malate!
- Beta-Alanine & Histidine: The Muscle Burning Fire Fighters!
Needless to say, I am completely psyched about a new compound I wish to introduce you to today - Orotic Acid (OA). So pay close attention, because OA is another performance enhancement breakthrough that is going to blow up the spot and take the sports nutrition world by storm in 2005.
Obviously I wouldn't be doing my job if I didn't give you an OA break-down. So without further ado, let's begin a brief synopsis on the next big thing in 2005 - Orotic Acid! OA is an intermediate in the pyrimidine pathway (see figure 1.) and is found in high concentrations in cow's milk.

Pyrimidine nucleotides are important constituents of phospholipids (structural components) in cell membranes such as the sarcolemma of muscle tissue. These nucleotides are also important constituents of RNA, the building blocks of new muscle and other cellular tissues.
There have been a variety of studies from the USSR by Meerson et al.[1, 2] in the 1960's showing the potential of this compound as an ergogenic aid to exercise. But that is old-school research, what is up with the new-school studies you ask?
In this, the first complete article covering this truly remarkable new supplement, we hope reveal its genuine potential in enhancing your athletic performance and body composition.

Transfer, Synthetic And Resynthesis Pathways
Transfer and synthesis are two of the key principles to understanding the mechanisms behind energy production in muscle. By truly understanding this mechanism we can optimize performance.
The key / primary energy source for our bodies is ATP (adenosine triphosphate) and its transfer and or activity in muscle is fast and furious. Only from the energy released by the breakdown of this compound can the cells perform work. Synthetic systems are pathways where the energy ATP or similar energy releases are maintained through compounds that assist in their regeneration.
|
 |
|||

When transfer rates are high we tend to see synthetic pathways (i.e. pyrimidine or purine nucleotide; see figure 1) are low.
An example of this is can be seen by looking at muscle tissues where we find high ATP transfer rates yet purine synthetic rates are low (Rossi 1975) [3]. Although the concentration of high-energy compounds such as ATP are low in places were transfer is high, the rates of resynthesis are just the opposite.
An great example of this can be seen in the heart (myocardium) were uracil and cytosine (pyrimidine bases) have turnovers of about 7 and 14% per hour with adenine nucleotiodes such as ATP having a turnover of only 2% [3]. So how does OA act as a precursor to pyrimidine base synthesis and how can it aid ATP production and maintenance?
Orotic Acid can maintain ATP in 3 different ways and more ATP means better performance:
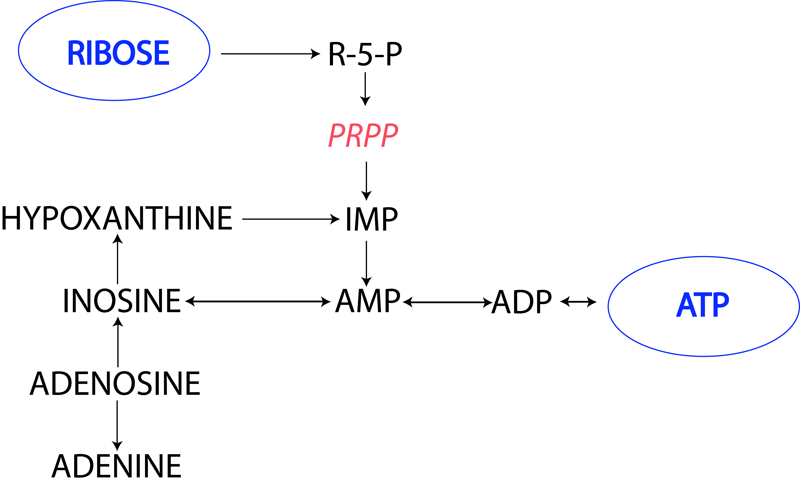
- The production of ribose moieties (R-1-P and R-5-P) for the salvage or de novo synthesis of purine nucleotides by a process know as "ribose transfer" [4]. Many of you may remember ribose (a sugar component of ATP) supplementation being touted as an enhancer of high intensity repetitive exercise performance. [5]. for a quick recap on the energy producing pathways of ribose see figure 2.
The way OA enhances the formation of ATP is by providing the direct precursor to phosphoribosylpyrophosphate (PRPP) a key energy-supplying compound in the de-novo synthesis of purine and pyrimidine nucleotides (I know that's a real mouthful). The reserve of PRPP is poor because of the low capacity of the pentose phosphate pathway. Therefore the generation of ribose-5-phosphate (R-5-P) the immediate precursor to PRPP ribose supply can be enhanced with OA and as a result greater ATP regeneration (See figure's 1 & 2).
- The second pathway is by increasing the pool of Uridine Monophosphate (UMP), which requires ATP to convert to further downstream pyrimidine nucleotides (UTP & CTP) (See figure 1 & 3). By increasing the levels of these nucleotides the draw on ATP reserves to synthesize them is decreased thereby enhancing the availability of ATP for other metabolic process such as energy and contractile producing pathways [6].
- The final pathway is by means of elevation of intracellular glycogen stores. These elevated glycogen stores generated from a greater uptake of glucose lead to an enhanced capacity for ATP resynthesis from anaerobic glycolysis (Kypson et al. 1978) [7].
So, what is the direct evidence for the influence of OA as an enhancer of ATP?
As of now there are 2 very important studies looking at the influence of OA on ATP levels. Richards et al. (1997) [4] continued on from the hypotheses set by Williams et al. (1976) [8] and Donohoe et al. (1993) [9] that orotic acid may provide intermediates that can be used in the synthesis of ATP.
Following a 2 day loading phase with orotic acid measurements of total adenine (ATP, ADP, and AMP) a pyrimidine nucleotides (cytosine and uridine) were taken with a resultant 121% rise is muscle tissue nucleotides. Further work substantiated the influence of orotic acid supplementation on ATP [10].
So we have proven the importance of OA in maintaining ATP levels in muscle tissues, but what does this exactly mean to us athletes in the trenches? Increased muscle demand for adenosine triphosphate (ATP) can decrease the total adenine nucleotide (TAN = ATP, ADP, and AMP) and total pyrimidine (TPN = cytosine, uridine and there phosphate bound metabolites) pools.
These metabolic reservoirs may be significantly decreased as a result of myocardial or skeletal muscle ischemia (an inadequate supply of blood), [11] certain metabolic diseases, [12] but of most importance to the us athletes trying to reach the next level is during repeated intense skeletal muscle contractions or in repetitive high-intensity exercise [13].
So prolonged and /or repetitive depletive stress placed on the adenine nucleotide pool may result in a greater reliance on the supply of ATP precursors "pyrimidine pool".
In English; depletion of the pyrimidine pool that occurs with intense exercise may decrease the energy generating capacity of muscle tissue [13].
So far so good however I bet most of you are asking the question of what information on humans exists on exercise performance and its proven beneficial effects following OA (orotic acid) intake.
Orotic Acid & Carnoside Synthesis
Carnosine's influence on controlling pH (Acidity in muscle) and the associated extension of high intensity exercise performance has been well described [14]. During exercise excessive lactate production is part of the extreme fatigue process; however it is the protons produced at the same time that restricts further performance.
To date ß-alanine has been shown as the limiting factor for the enhancement of muscle carnosine [15] concentrations. However, based on previously published work this may not be the only compound that can enhance cellular carnosine stores [6] that lead to an enhancement in performance of high intensity repetitive bouts of exercise.
The first thing we need to understand about OA and carnosine concentrations is why another nutrient outside of ß-alanine (already shown to be highly effective in elevating muscle carnosine) would be effective or useful in elevating muscle carnosine when you could just use Beta-Alanine?
Make no mistake about it Beta-Alanine it is the real deal in terms of increasing explosive power and muscle carnosine stores, but there's more... ß-alanine is not only used to synthesize carnosine it is also degraded to RNA for protein synthesis and cellular repair [6]. So for athletes who require increased synthesis of proteins to repair damaged muscle tissues there is a greater drain on RNA meaning a greater use of ß-alanine meaning less available for carnosine synthesis.
One way of looking at the proportion of ß-alanine used for synthesizing RNA is to inhibit RNA synthesis. The injection of 6-azauracil a potent inhibitor of RNA synthesis given to rats allows ß-alanine to be used primarily in the synthesis of carnosine.
These studies show following the inhibition of RNA synthesis a huge increase in ß-alanine uptake (70%) and a greater elevation in carnosine synthesis [6]. This shows that a large proportion of ß-alanine is indeed used for RNA synthesis and growth, and that ß-alanine is truly a multifunctional nutrient outside of its currently defined role as a precursor for carnosine synthesis. That's why, the use of compounds that can directly support ß-alanine formation will increase its availability for carnosine synthesis in times of elevated growth and cellular regeneration.
Aonuma's group were pioneers in the field identifying the metabolic precursors of carnosine and work in the 1960's provided great insights into what elevates carnosine stores and what does not.
As a result of their investigative work a clear pathway was identified for the conversion of orotate (OA) to ß-alanine and carnosine in skeletal muscle tissues. A detailed synthetic pathway of carnosine synthesis can be seen in figure 3.

Orotic Acid & Exercise Capacity
Two great studies show the real influence of orotic acid supplementation given orally on exercise capacity and biochemical markers that influence muscle growth (i.e. cortisol) and recovery (Leukocytes). Geiss et al. (1998) [16] studied a group of patients with coronary heart disease before and following 4 weeks of 3g per day of magnesium orotate supplementation. Following the 4 weeks of orotate therapy exercise duration was significantly increased by a massive 21%.
In the second human study we get our first glimpse of the real power of orotic acid on human performance in a healthy population "triathletes" [17].
In a double blind randomized trial (researchers and subjects did not know who was on supplement or placebo) carried out again over 4 weeks with or without 3 grams per day of magnesium orotate assessed swimming, running and cycling performance. At the end of the supplementation period all exercise assessments were improved (faster) with markers of stress (leukocyte count and cortisol) significantly lower in the orotate group [17].
This study provides us with the first real evidence that orally consumed orotate enhances exercise performance. Whether by enhancement of muscle glycogen, ATP levels, muscle carnosine, mineral status or a combination of all 4 is unknown. However the take home message of course remains the fact that orotic acid WORKS!
Other studies from back in the early 60's give evidence of other functional effects based on animal models including accelerated skeletal muscle hypertrophy (muscle growth) following supplemental orotic acid [1, 2]. work by the Meerson group demonstrated enhanced swimming performance as a function of muscle hypertrophy and energy supply [1, 2] following orotate supplementation.
More recent work on isolated human muscle fibres has shown that elevation of pyrimidine base concentration (Uridine = elevated following orotate feeding in humans and mice) enhances the contractile capacity of muscle tissue [18].
This suggests that there maybe a mechanism for enhanced force production following orotate supplementation as well as its many other beneficial effects. This is good news for those powerlifters out there!
Ribose + Nucleotides: Not An Additive Combination
As we discussed earlier ribose supplementation positively enhances exercise performance by aiding the resynthesis of ATP (see figure 2). Therefore, it is possible that the combination of ribose and other purine and pyrimidine precursors i.e. adenine, hypoxanthine, and orotic acid could have additive if not synergistic effects on energy metabolism if not performance. However, because orotate can increase ribose stores taken alone; studies by Munsch et al. (1992) [19] proved no additive influence of combining these nutrients on functional recovery.
Basically beware of dietary supplements that claim a ribose and orotate combo is more beneficial then orotate alone as it seems orotate may provide benefits as great and beyond that ribose. Again, this is still relatively new information to athletes and I expect many supplement companies to include OA, although they may lack the fundamental understanding of how it works and what is best to use in conjunction with it.
Safety Of Orotate Supplementation
The build up of pyrimidine nucleotide precursors in the liver and the subsequent increase of very-low-density-lipoproteins, VLDL (fats) in the rats has been a concern to many regarding supplement orotate use in humans!
However, based on all available evidence (including human studies) this is not a concern and seems to be a condition confined to only to rats for many reasons including: Low levels of 5 nucleotidase activity (not low in comparison in humans) [20], very low metabolism of OA by other tissues outside of liver and kidney [21], and low excretion rates from bile and urine [22, 23].
|
|
|||

Because of these poor metabolic handling problems in rodents a slow but steady build up of plasma uridine and cytodine occur and soon a subsequent accumulation of VLDL in said tissues [24]. These metabolic inadequacies in rodent tissues are not present in the human body. So based on the current evidence there seems to be no negatives of orotate supplementation and those suggesting other wise clearly have misunderstood the biochemistry behind orotate supplementation.
Furthermore in the only well controlled human studies carried out on patients with clinically diagnose heart disease as well as healthy athletes show only positive effects of orotate supplementation on both heart function, blood profiling (lipid profiling) and exercise performance/capacity [16, 17].
Further data on supplementation on hamsters, mice and guinea pigs with OA show no influence on lipid metabolism suggesting that the negative of OA are truly specific to rats [4, 20-24].
So, What Is The Bottom Line?
OA seems to be one of the most effective and exciting new nutrients that is under studies in the field of sports nutrition. Its future as an effective addition to dietary interventions look to go from strength to strength based on current data. So what might be its potential uses and additive combinations as a dietary supplement?
One possible use is in combination with creatine the undisputed heavy weight champion of supplements in human sports nutrition.
Creatine is a vital component for the regeneration and maintenance of ATP during intense muscular contractions its influence on performance enhancement has been shown time and time again in peer reviewed studies [25, 26]. Work by Rosenfeldt et al. (1998) [27] showed under hypoxic (lack of oxygen) conditions (found following anaerobic exercise) elevation of intramuscular uridine levels increased muscle phosphocreatine and creatine levels 38% and 3%, respectively.
This shows yet another great effect of orotic acid and suggests a possible synergism with supplemental creatine for enhancing muscle efficiency and power production as well as recovery between repetitive exercise. The most effective nutrients that seem already to work in synergy with orotate is magnesium. Magnesium a mineral found to be deficient in many of us [28] and increased losses through sweat and urinary outputs increases the incidence of deficiency in those exercising regularly.
In closing OA seems to have all the necessary qualifications behind it to improve exercise capacity and in turn physique change. I am sure over time this nutrient will become widely used as a natural ergogenic aid that can safely enhance your performance and your body but has also wider application in the medicinal community.
Be on the lookout for Orotic Acid to be featured in our revolutionary new creatine matrix CLOUT.
The Final Recap!
The Main Mechanisms Of Orotic Acid:
- Enhancement and maintenance of ATP pools
- Increased glucose uptake.
- Increased ribose moieties (R-1-P & R-5-P) leading to enhance ribose formation.
- Increased muscle carnosine stores.
- Support and enhancement of muscle hypertrophy (via RNA synthesis support during growth).
- Elevates uridine a DNA base shown to increase muscle contractile capacity.
References
- Meerson FZ. (1969) The myocardium in hyperfunction, hypertrophy and heart failure. Circ.Res. 24/25 (Suppl.2): II146 - II155.
- Meerson FZ, Rosanova LS. (1967) Effect of 2703 actinomycin and combination of nucleic acid synthesis activators on the development of fatigue and fitness. Doki. Akad. Nauk. SSSR: 166: 496-499.
- Rossi A. (1975). 32-P-labelling of the nucleotides in position in the rabbit heart. J .Mol Cell. Cardiol. 7: 891-907.
- Richards SM, Conyers RAJ, Fisher JL, Rosenfeldt FL. (1997) Cardioprotection by Orotic Acid: Metabolism and mechanism of action. J.Mol.Cell.Cardiol. 29: 3239-3250.
- Kreider RB, Melton C, Greenwood M, Rasmussen C, Lundberg J, Earnest C, Almada A. (2003) Effects of oral D-ribose supplementation on anaerobic capacity and selected metabolic markers in healthy males. Int.J.Sport.Nutr.Exerc.Metab. 13: 76-86.
- Aonuma S, Hama T, Tamaki N, Okumura H. (1969) Orotate as a -alanine donor for anserine and carnosine biosynthesis and effects of actinomycin D and azauracil on their pathway. J.Biochem. 66: 123-133.
- Kypson J, Hait G, Mathew R. (1978) Effects of uridine on the performance and the metabolism of oxygenated and hypoxic rabbit hearts. J Mol Cell Cardiol. 10: 545-565.
- Williams JF, Donohoe J, Lykke A, Kolos G. (1976) Studies using orotic acid for improving the controlled development of myocardial hypertrophy. Aust.NZ.J.Med. 6 (Suppl.2): 60-71.
- Donohoe JA, Rosenfeldt FL, Munsch CM, Williams JF. (1993) The effect of orotic acid treatment on the energy and carbohydrate metabolism of the hypertrophing rat heart. Int.J.Biochem. 25: 163-182.
- Olivares J, Rossi A. (1982) Incorporation of orotic acid in myocardial uridine nucleotides: effect of isoproterenol and ribose. J.Physiol. (Paris). 78: 175-178.
- Newman MAJ, Chen X-Z, Rabinov M, Williams JF, Rosenfeldt FL. (1989) Sensitivity of the recently infracted heart to cardioplegic arrest. J. Thorac .Cardiovasc. Surg. 97: 593-604.
- Sumi S, Kidouchi K, Imaeda M, Asai M, Ito T, Wada Y. 1997. Urinary orotic acid in healthy adults and patients with various diseases. Clinica.Chemica.Acta. 266: 195-197.
- Yamamoto T, Moriwaki Y, Takahashi S, Tsutsumi Z, Yamakita J, Higashino K. (1997) Effect of muscular exercise on the concentration of uridine and purine bases in plasma ATP consumption induced pyrimidine degradation. Metabolism. 46:1339-1342. 14. Harris RC, Dunnett M, Greenhaff PL. (1998) Carnosine and Taurine contents in individual fibres of human vastus lateralis muscle. J.Sport.Sci. 16: 639-643.
- Harris RC, Hill C, Wise J, Tallon MJ. (2003) Effect of combined b-alanine and creatine monohydrate supplementation on exercise performance. Med.Sci.Sport.Ex. 35 (5): abstract 1206.
- Geiss KR, Stergiou N, Neuenfeld HU, Jester HG. (1998) Effects of magnesium orotate on exercise tolerance in patients with coronary heart disease. Cardio.Drug.Ther. 12: 153-156.
- Golf SW, Bender S, Gruttner. (1998) On the significance of magnesium in extreme physical stress. Cardio.Drug.Ther. 12: 197-202.
- Vianna-Jorge R, Oliveira CF, Mounier Y, Suarez-Kurtz G. (1998) Functional effects of uridine triphosphate on human skinned skeletal muscle fibers. Can. J. Physiol. Pharmacol. 76: 110-117.
- Munsch CM, Williams JF, Conyers RAJ, O'Halloron K, Langley LH, Rosenfeldt FL. The effect of orotic acid and ribose on the impaired tolerance of the recently infracted rat heart to cardioplegic arrest. In: Williams JF, ed. Orotic acid in cardiology. New York: Thieme Medical, 1992; 46-58.
- Bontempts F, Vincent MF, van den Berghe G. (1993) Mechanisms of elevation of adenosine concentrations in anoxic hepatocytes. Biochem.J. 290: 671-677.
- Gasser T, Moyer JD, Handschumacher RE. (1981) Novel single-pass exchange of circulating urindine in rat liver. Science. 213: 777-778
- Fausto N. (1972) The conversion of orotic acid into uridine 5-monophosphate b y isolated perfused normal and regenerating rat livers. Biochem.J. 129: 811-820.
- Handschumacher RE, Coleridge J. (1979) Hepatic and biliary transport of orotate and its metabolic consequences. Biochem.Pharmacol. 28: 1977-1981.
- Pels Rijcken WR, Hooghwinkle GJM, Ferwerda W. (1990) Pyrimidine metabolism and sugar nucleotide synthesis in rat liver. Biochem.J. 266:777 - 783.
- Greenhaff PL, Casey A, Short AH, Harris R, Soderlund K, Hultman E. (1993) Influence of oral creatine supplementation of muscle torque during repeated bouts of maximal voluntary exercise in man. Clin.Sci. (Lond). 84: 565-71.
- Harris RC, Soderlund K, Hultman E. (1992) Elevation of creatine in resting and exercised muscle of normal subjects by creatine supplementation. Clin.Sci. (Lond). 83: 367-374.
- Rosenfeldt FL, Richards SM, Lin Z, Pepe S, Conyers RAJ. (1998) Mechanism of cardioprotective effect of orotic acid. Cardio.Drug.Ther. 12: 159-170.
- Gold SW. Is magnesium a limiting factor in competitive exercise? A summary of relevant scientific data. In: Gold SW, Dralle D, Vecchiet L. ed. Magnesium 1993. London: John Libbey, 1994: 209-219.