
Protein and Lipogenesis are a couple of words that for all bodybuilders and fitness enthusiasts sound like words that do not match.
How many times have we heard from our Physician, from the dietician, from medical TV shows that to get a better body we need to negatively shift the daily energetic equilibrium by increasing the daily calorie expenditure and/or our daily caloric intake so that it is lower than the expenditure?
How many times have we heard from trustworthy health professionals such as nutritionists, who have a wide influence on the public, that all the caloric excesses get transformed into triacylglicerols and then stored as fat (subcutaneous adipose tissue)?
And how many other times have we heard (but not believed) that also protein (if consumed in excess) may trigger the lipids biosynthesis in the same manner and at the same rate as carbohydrates and fats do when consumed in excess than the daily requirements?
If it's true, how do you explain the excellent muscular definition and the very chiselled abdominal wall of bodybuilders, athletes and fitness enthusiasts whose diets are very similar - even though there are some differences - with most of them using the high protein and low carbohydrate regimen for achieving muscular definition?
 Furthermore, how so you explain the high muscular quality induced by a high protein diet, independently from caloric intake?
Furthermore, how so you explain the high muscular quality induced by a high protein diet, independently from caloric intake?
Here it's obvious that arithmetic is not clear at all, because the evidence of the facts don't support the theory of the over-simplified mathematic count as the determinant factor that influence body composition (Body composition is the result of Fat Free Mass to Fat Mass ratio).
This is explained by the fact that our organism is much more sensible to an "hormonal and biochemical mathematic" rather then the merely algebraic mathematic.
Here is important to assert, and it's even evident, that the single individual cannot be considered like a calorimetric bomb (for which 1 gram of carbohydrate is not different from a gram of protein since they do have the same caloric output: 4kcal per gram) since it -the calorimetric bomb- goes far beyond from any endocrine and biochemical consideration.
In fact we're right Dr. Barry Sears with its "Enter the Zone", Dr. Mauro Di Pasquale with its "The Metabolic Diet" and Dr. Atkins with its "Atkins Diet": On a isocaloric diet, a high-protein diet is more lipolytic and less lipogenetic that the high-carb diet.
Experts on the matter such as Will Brink and Mauro Di Pasquale state the protein molecule is the macronutrient that is less likely to be converted into triglycerides. So, why does all this discrepancy exist within the medical community on this matter?
In order to make clarity on the reasons that lie behind such statements- clearly conflicting one to another- the biochemistry is the main subject to which is necessary to refer to fully understand this concern.
Starting from the biochemical analysis of the fatty acids biosynthetic events, we will see what conflicting opinions have in common and share the same biochemical steps, but different is their interpretation.

Biochemistry Of Fatty Acids Biosynthesis
The biosynthesis of fatty acids occurs starting from its immediate precursor, Acetyl-CoA, in the citosol of hepatic (liver) cells, of adipose tissue, and mammal glands of superior animals.
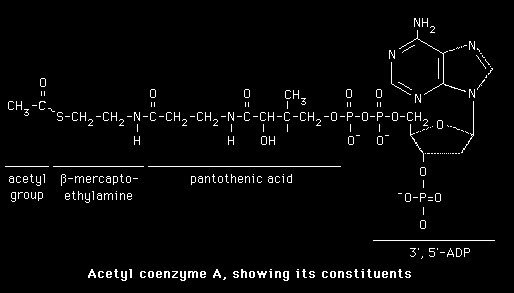
Acetyl-CoA, "Primer" molecule of fatty acids synthesis, derives from glucose, fatty acids, and amino acids catabolism. This molecule (Acetyl-CoA) may face different destinies according to the cellular energetic conditions.
If conditions of low cellular energetic charge occur, Acetyl-CoA gets oxidised inside the Krebs Cycle (a.k.a "Citric Acid Cycle" or "Tricarbossilic Acid Cycle") to extract energy through GTP production as well as the reduction of NAD+ and FAD+ into NADH and FADH respectively, which will cede their electrons to the "Electrons Transport Chain" in order to promote the processes of "Oxidative Phosphorylation."
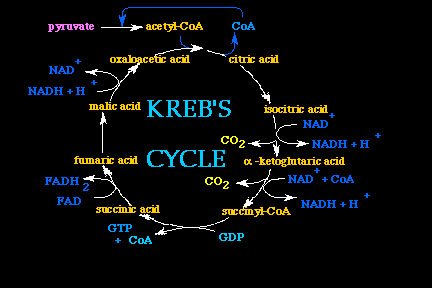
On the other hand, instead, when the cellular energetic charge is high, the processes for energy extraction get substituted by the biosynthesis processes, which implys (in the case of fatty acids biosynthesis) the non-oxidation of Acetyl-CoA but rather, its use as "Primer" substrate for the upcoming fatty acid lengthening.
Furthermore, the presence of the Citrate is necessary for the fatty acid biosynthesis to take place. The fatty acid biosynthesis moves toward the formation of Palmitic Acid which has 16 carbon atoms.

Palmitic Acid Molecule
The fatty acid biosynthesis is catalyzed by a series of proteic enzymes named "Fatty Acid Synthetase Complex". Of the eight bi-carbons units of Acetyl-CoA necessary for the synthesis of Palmitic Acid (16 carbon atoms), only one of them is supplied by Acetyl-CoA itself, since the other 7 come under the form of Malonil-CoA.
The biosynthesis reduction power is supplied by NADPH.
Before I go deeper in the detail of the biochemical analysis of the processes at the base of fatty acids synthesis, it is important to keep in mind that in order for the biosynthesis process to occur, the presence of Acetyl-CoA, Citrate, and NADPH are required together.
As already afore mentioned, the fatty acids biochemical processes occur in the citosol therefore implying that Acetyl-CoA becomes inactive during the Krebs Cycle that occurs inside the Mitochondria.
Acetyl-CoA is not able to pass from the mitochondria to the citosol (the place of biosynthesis). Instead the Citrate formed inside the Krebs Cycle (inside the mitochondria) by the reaction of Acetyl-CoA and Oxaloacetate, can pass through mitochondrial membrane to the citosol by means of the "Tricarbossilic Acids Transport System".
Inside the citosol, Acetyl-CoA get reformed by the "ATP-Citric Lyase enzyme" that breaks down the Citrate. Previously mentioned that along with the Primer (Acetyl-CoA), essential to trigger the biosynthesis, the other 7 bi-carbon units that complete the synthesis must reach the "Fatty Acid Synthetase Complex" under the form of Malonil-CoA.
Malonil-CoA gets formed by the union of one molecule of Acetyl-CoA and a molecule of CO2. This reaction is catalyzed by the "Acetyl-CoA Carboxylase enzyme" which contains Biotin (also known as Vitamin H), which in turn serves as an intermediate carrier of a CO2 molecule.
Acetyl-CoA Carboxylase is an allosteric enzyme which catalyse a reaction that constitutes the main step that regulates or limits the speed of fatty acid biosynthesis.
This enzyme is inactive without the presence of its positive modulators: Citrate and Isocitrate- therefore the remarkable allosteric stimulation of this enzyme induced by the presence of the Citrate explains why the Citrate itself is required for the fatty acid synthesis.
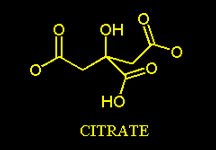
The brief review of the biochemical events so far discussed, clarify what motivations are at the base of the common belief that has always induced the medical community to support the theory that all the energetic excesses, independently on how the macronutrients are shared among themselves (their ratio within a given meal) and their percentage in a given daily caloric intake, lead to the accumulation of the excess energies (calories) as triglycerides in the adipose tissue (Fat tissue) since the catabolism of glucose, amino acids and fatty acids moves toward a common pathway : Acetyl-CoA!
Basically this argument would lead everyone who have studied biochemistry to conclude that also proteins are able to generate lipogenesis if the upcoming described following factors wouldn't be taken into consideration:
- A) The endocrine response of Insulin-Glucagon induced by the food consumption; and
- B) The influence that the elicited hormonal output has on the destiny of the biochemical reactions that lead to the biosynthetic events.
4 Points That Greatly Influence Lipogenetic Events
Point 1
The Acetyl-CoA formed by the elementary macronutrients catabolism enter the Krebs Cycle and reacts with the Oxaloacetate to form Citrate. As afore discussed, the Citrate (in order to trigger the lipogenetic processes) has to exit the Krebs Cycle (which occurs inside the mitochondrion) to move toward the citosol (the place of biosynthesis). The exiting of Citrate from the Krebs Cycle is induced by insulin!
Once in the citosol, Citrate, gets broken down and Acetyl-CoA (which acts as the Primer) gets reformed again. Previously has been also said that the lengthening of the Palmitic Acid (fatty acid) is possible by the formation of the Malonil CoA which is formed by the reaction between Acetyl-CoA and CO2 and that this reaction is catalyzed by the Acetyl-CoA Carboxylase enzyme.
Acetyl-CoA Carboxylase is not stimulated by the Citrate presence only, but by the insulin as well! Therefore on a low-carb nutrition program, the insulinemic response is not as equal as the one seen while on a high-carb diet (60% carb or more) although the calories between the two type of diets are the same.
Furthermore, also the exiting of the Citrate from the Krebs Cycle and the activation of Acetyl-CoA Carboxylase result both to be altered by the variation of the insulinemic levels rather than the caloric intake itself.
This consideration, favourable for the poor insulin stimulation, could even be turned in a more profitable condition if the carb intake comes from low glycemic index carbohydrates sources.
So, the low-carb intake plus the low glycemic index of the carbs make turn out a low "Glycemic Load" which is the key element in the stimulation of the insulin incretion.
What is really important is to keep in mind that the main responsible for eliciting the insulin output is the Glycemic Load as the result of the glycemic index of the carb ingested times the carb content of the food rather than the glycemic index itself.
While glicemic index only measures how quick a kind of carbohydrate triggers the rise in glycemia (circulating blood sugar), on the other hand Glycemic Load gives a fuller picture because it takes into consideration both key factors: food carb-content and Glycemic Index.
These two information together allow to access to the real food's effect on blood sugar.
For example:
- 100gr of watermelon contains 3.7gr carbs with a 72±13 glycemic index
- Glycemic Load= (3.7 x 72±13/100)= 2.2 to 3.1
100gr dried apples contain 92.2gr carbs with a 29±5 glycemic index
Glycemic Load =(92.2 x 29±5/100)= 22.1 to 31.3
As a matter of fact watermelon has a far greater Glycemic Index than dried apples, but it's Glycemic Load is far lower than dried apples.
A Glycemic Load of 20 or more is high, between 11 to 19 inclusive is medium and a Glycemic Load of 10 or less is low.
Point 2
Furthermore, when the cellular energetic charge is low or in those cases when the glycemia is low for a momentary deficit (as in those cases induced by the training) or protracted (as in the case of low-carb diet) gluconeogenesys processes get activated.
One of the substrates used for glucose production (when glucose is not introduced through the diet or administered through other exogenous sources) is an Intermediate of the Tricarbossilic Acid Cycle (a.k.a. Krebs Cycle): Oxaloacetate.
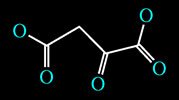
Oxaloacetate Molecule
Gluconeogenesys processes occur in the liver.
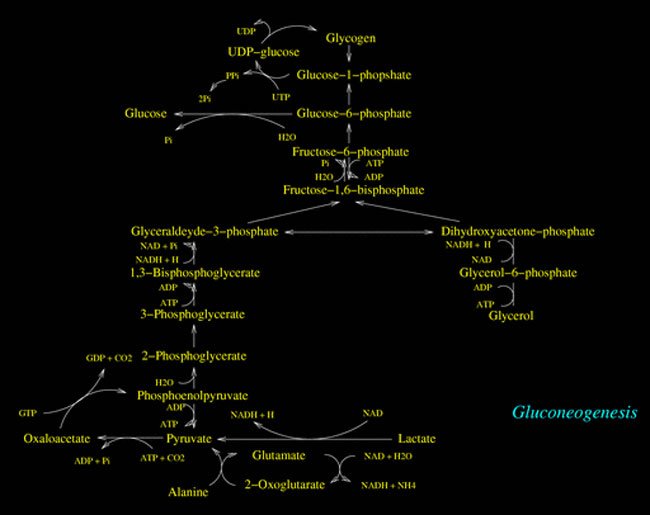
When Oxaloacetate is used as glucose precursor, it gets subtracted from the Krebs Cycle thus inhibiting the Acetyl-CoA (which comes from the nutrients' catabolism) to enter the Krebs Cycle and consequently not able to react with Oxaloacetate for the upcoming Citrate formation.
The absence of this reaction (Oxaloacetate+Acetyl-CoA=Citrate) turns out not only in a lesser bioavailability in the citosol of Acetyl-CoA (the molecule which triggers the start of biosynthesis processes) which not being able to enter the Krebs Cycle goes toward the Keton Bodies formation (which will be in part degraded for energetic purposes by the metabolically active tissues -skeletal muscle- and in part excreted through respiration and urines), but also in a lacking of Citrate in the citosol (as afore mentioned, if present in the citosol it would act as a positive modulator for the allosteric stimulation of the enzyme that catalyze the Malonil CoA formation: the Acetyl-CoA Carboxylase).
Point 3
The Insulin-Glucagon Ratio after consuming a high protein meal. After consuming a carbohydrate-containing meal, ematic glucose (glycemia) arises, insulin levels increase and simultaneously inhibit Glucagon secretion.
Instead, after consuming a protein-containing meal, aminoacidemia increases which triggers the insulinemic incretion as well. However the plasmatic concentration of glucose cannot be sustained because there's not any glucose intake with the consumption of the protein meal. As consequence, Glucagon incretion increases too so that glucose plasma concentration is supported by hepatic glycogenolisys and gluconeogenesys too.
Glucagon is a polypeptide hormone which acts on the liver - promoting hepatic glycogenolisys - and on the adipose tissue as well- inducing lipolisys (the triglyceride break down process).
The lipolisys process induced by glucagon is justified also by the fact that the triglycerides catabolism leads to the detachment of glycerol from the triglyceride molecule (which is formed by 3 fatty acids molecules and one glycerol molecule) which is an important substrate for gluconeogenesys processes.
Therefore the Insulin and Glucagon output is strictly dependent by the quality of the foods ingested and not by the over-simplified mathematic counting of calories independently of the food sources where the nutrients come from.
Point 4
Oxidative amino acids degradation.
It is well known that amino acids are the elementary units for protein synthesis. Furthermore they have many other functions. They are precursors of other important bio-molecules such as: hormones, purines, pyrimidines, porfirines and some vitamins too.
Nevertheless, in the case their intake through the diet exceed the quantity required for the normal protein turn-over, they are used as an energy source. To be used as fuel for energy, they lose their amminic groups and their carbon skeleton undergo two main processes: 1) glucose conversion in the gluconeogenesys process -OR- 2)oxidation into CO2 inside the Krebs Cycle.
The removal of the amminic group is a fundamental step in the amino acid catabolism and there are two enzymatic ways that lead to its removal: Transamination and Oxidative Deamination.
In the transamination phase the amminic group is transferred to the carbon atom of a Alpha Keto-Acid which is in most of the cases the Alpha Ketoglutaric Acid and transforming it through the ammination process into L-Glutammic Acid.
The L-Glutammic Acid formed by the transaminase action may undergo a rapid oxidative deamination catalyzed by Glutammic Dehydrogenase and therefore losing, as NH4+, the amminic groups derived from other amino acids.
For this event to occur a Dehydrogenation process is required, and the Glutammic Dehydrogenase may use either NAD+ and NADP+ as electron acceptors, but it prefers NAD+.
The NADH formed then gets oxidised in the "Electrons Transport Chain".
This is an important step because the preference of NAD+ rather than NADP+ by the Glutammic Dehydrogenase turns in a lesser presence of the molecule that brings reduction power (NADPH) which is essential for the formation and lengthening of the fatty acid chain.
All this detailed biochemical explanation is not intended to state that all the energetic excesses that come from excess protein consumption don't induce lipogenesis at all, since the 2nd law of thermodynamics is still valid.
But all this complex scientific description is important to point out that the different ratios among macronutrients within the same caloric intake generate different responses because different are the hormonal output induced by the nutrients and therefore different will result the ways and the measure by which the energetic excesses get transformed into subcutaneous body fat.
Such differences in hormonal responses which are elicited by the introduction (consumption) of different chemical molecules (carbohydrates, proteins, lipids) generate even more significant changes in body composition when all this refers to an hypocaloric regimen.
In fact the "Landmark Study" of Kewick and Pawan represents the scientific proof of previously mentioned. The study submitted a group of people to a hi-fat low calorie (1000kcal) regimen. The patients of his study decreased significantly their bodyweight.
When the same group of patients underwent through the same low calorie regimen (1000kcal) but different from the former for its hi-carb content, there was not any decrease in bodyweight (Kewick A. and G.L.S. Pawan.- "Metabolic study in human obesity with isocaloric diets high in fat, protein or carbohydrate", in "Metabolism", 1957, 6:447-460).
Conclusions
The mathematic caloric count gives us information about our daily energetic balance, on the other hand, it's not able itself to supply unambiguous explanations about the body composition changes.
All this means that the caloric counting itself has not to be taken into consideration as the only key factor to planning a nutrition program when the aimed and desired goal is the improvement of body composition which results more linked to the shift of the Glucagon-Insulin ratio (in favour of the former) rather than the total sum of the energetic values of the nutrients introduced through the diet.
As you all may notice, biochemistry and endocrinology knowledge are the tools to possess to better understand and to manipulate body composition through dieting.
Whatever the level of body shape you want to drive your body to, do it successfully according to science!
Keep Growing And Stay Healthy!